
Introduction
All members of human societies are characterized by a deep concern over issues of morality, justice, and fairness (Decety & Yoder, 2017). In fact, humans are unique among other living creatures in establishing cultural organizations to enforce particular social norms, including institutions designed to evaluate the acceptability of individuals’ behaviors and assign appropriate punishments to those who violate particular norms (Buckholtz & Marois, 2012). Regardless of how morality is conceptualized, many scholars, following Darwin (1871), have made the claim that it is an evolved aspect of human nature that facilitates cooperation in large groups of unrelated individuals (Tomasello & Vaish, 2013). Associating in groups improves the chances of survival compared with solitary existence. Moral norms provide safeguards against infringements on safety or health, and reinforcement of moral behaviors minimizes criminal behavior and social conflict. In these ways, morality makes human society possible.
There are at least two reasons to think morality bears the imprint of natural selection. While nonhuman animals obviously don’t reason explicitly about right and wrong, good and bad, just and unjust, or vice and virtue, some exhibit behaviors which seem to incorporate elements of human morality. Many species cooperate, help their kin, and care for their offspring (Tremblay, Sharika, & Platt, 2017), and some manifest inequity aversion (Decety & Yoder, 2017. Likewise, while socialization influences moral development and explains why moral rules change with space and historical time, human infants enter the world equipped with cognitions and motivations that incline them to be moral and prosocial (Hamlin, 2015). Such early emerging predispositions toward prosocial behavior, and sociomoral evaluation reflect prewired capacities that were adaptive to our forebears. Members of homo sapiens cooperate with and help non-relatives at cost to themselves at a rate that is unmatched in the rest of the animal kingdom. This penchant for cooperation with unrelated individuals explains why our species successfully colonized the entire planet (Marean, 2015). However, this does not imply that morality is itself an adaptation favored by natural selection. Instead, the moral sense observed in humans may be a consequence of several cognitive, executive, and motivational capacities which are the attributes that natural selection directly promoted (Ayala, 2010).
Decades of research across multiple disciplines, including behavioral economics, developmental psychology, and social neuroscience, indicate that moral reasoning arises from complex social decision-making and involves both unconscious and deliberate processes which rely on several partially distinct dimensions, including intention understanding, harm aversion, reward and value coding, executive functioning, and rule learning (Decety & Cowell, 2017; Gray, Young, & Waytz, 2012; Krueger & Hoffman, 2016; Ruff & Fehr, 2014). Human moral decisions are governed by both statistical expectations (based on observed frequencies) about what others will do and normative beliefs about what others should do. These vary across different cultures and historical contexts, forming a continuum from social conventions to moral norms which typically concern harm to others.
In this article, we first discuss recent empirical progress in the neuroscience of social decision-making. Next, we examine the neural mechanisms underlying the components involved in morality. Then, a section is devoted to psychopathy because it constitutes a model for understanding the consequences arising from atypical neural processing and lack of concern for others and moral rules. Finally, we discuss how this social neuroscience perspective has valuable forensic implications, both in terms of understanding how jurors and judges make decisions about culpability and severity, and in predicting defendant’s future behavior. Understanding the psychological mechanisms and neurological underpinnings of how we make moral decisions sheds light on the diagnosis and treatment of the serious wrongdoers among us.
Social Behavior: An Overview
Social behaviors are behaviors that involve two or more organisms directly interacting in some way. While these kinds of behaviors are more common in some species than in others, almost all living things engage in a variety of them. Given there are generally a large number of members of a given species alive at a particular point in time, it is almost inevitable that different members will come to interact, and that selection pressures will result in the development of these interactions in an adaptive direction.
Because we rely heavily on social learning, human beings have developed the most extensive culture of any known species, and the richest matrix of social behaviors. We have constructed an extremely interdependent society, reliant on the division of labor and widespread, multi-level cooperation. Hence, being able to engage effectively in social interactions has become a prerequisite for succeeding as a human being, and human beings who suffer from social deficits are at a great disadvantage.
Social behavior is also very important for mice. While mice do not have the same level of culture as humans, they are highly social animals that tend to live in large groups. Common social behaviors between mice include mating, grooming, group sleeping, parenting, genital sniffing, biting, fighting, and squeaking. Most of these behaviors have analogues in humans, who also mate, communicate, parent and engage in aggression. The importance of sociality for mice allows them to serve as models for the research of human social disorders.
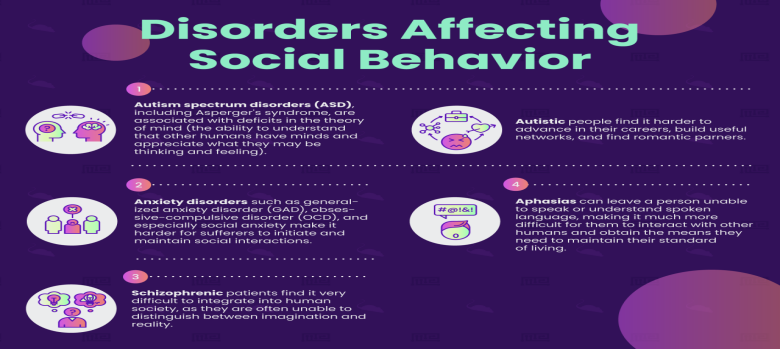
Disorders Affecting Social Behavior
In humans, there are many prevalent psychological and physiological disorders that have a detrimental impact on social behaviors. In fact, almost all serious disorders have some kind of impact on social behavior, given how central social interaction is to human life. In this section, we will discuss a few of these disorders. It should be noted that how psychological disorders are officially named and defined is subject to constant revision.

The Neuroscience of Social Behavior
An entire subfield of neuroscience, known as social neuroscience, is dedicated to understand how the brain produces and deals with social behaviors. As with all major behavioral areas, there is no one single part of the brain that is dedicated to social interaction, but rather a number of different brain regions that contribute to different aspects of the behaviors.
The Mirror Neuron System
Mirror neurons were first discovered in the brains of macaques;[7] there is evidence they are also present in the brains of humans and rats, as well as other creatures.[8][9] Mirror neurons are neurons that fire both when an animal performs a certain action, and when that animal observes the same action being performed by another individual. Hence, the neurons directly “mirror” the activity of another living system. Much speculation (often ungrounded) has been made about mirror neurons since their discovery, and it is important not to stray too far beyond what is scientifically known.
Imitating Behavior
Social learning is extremely important for human beings, perhaps more so than for any other species. Humans engage in many complicated daily activities—driving a car, using kitchen appliances, operating a computer and so on—that cannot be encoded in genetic instructions, and have to be learned through imitation. Humans appear to have a compulsion for social imitation, exemplified by children’s desire to copy whatever they see, and by adults doing things such as adopting accents and mannerisms. It could be that stereotyped practices are literally contagious, spreading through the human population via emulation in mirror neurons.
Social decision-making
Human lives consist of a constant stream of decisions and choices. Essentially, the study of decision-making attempts to understand the fundamental ability to process multiple alternatives and to choose an optimal course of action (Sanfey, 2007). This involves identifying rewarding stimuli to approach and aversive stimuli to avoid (Pessiglione & Delgado, 2015). However, successful decision-making also requires maintaining representations of short-term and long-term goals in order to maximize decision payoffs. Social decision-making specifically refers to decisions in which individuals consider and integrate the goals, beliefs, and intentions of other individuals into their decision-making calculus. Thus, social decision-making relies on Theory of Mind, the capacity to attribute mental states such as beliefs, intentions and desires to oneself and others. However, social decision-making is also at times constrained by social norms and associated punishments, which affect the expected value of specific response options.
The neural systems supporting social decision-making have been investigated from multiple perspectives, including neuroeconomics, cognitive neuroscience, and translational neuroscience (Figure 1). Converging evidence indicates that the components of social decision-making rely on the coordination of multiple neurocognitive systems which support domain-general processes such as stimulus valuation, perspective-taking, mental state understanding and response selection (Berridge & Kringelbach, 2015; Ruff & Fehr, 2014; Tremblay et a 2017).
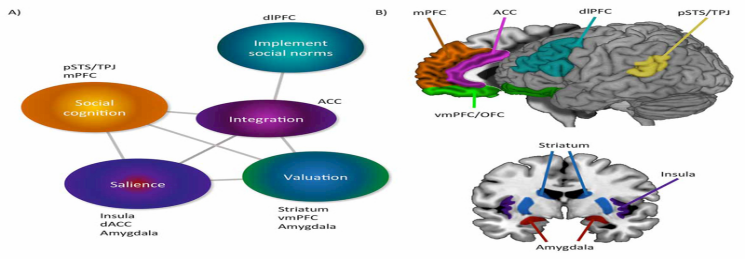
In non-social contexts, rewarding and aversive stimuli are associated with activity in largely separate neural networks (Pessiglione & Delgado, 2015). For instance, functional magnetic resonance imaging (fMRI) studies, have shown that the striatum, rostral anterior cingulate cortex (rACC), and ventromedial prefrontal cortex (vmPFC) are reliably recruited in response to rewarding stimuli. In contrast, dorsal ACC (dACC), anterior insula (aINS), and amygdala are more active in response to aversive stimuli including pain. More specifically, experiencing (as opposed to anticipating) a rewarding stimulus (e.g. eating ice cream or winning the lottery) is associated with neural responses in orbitofrontal cortex (OFC), ventromedial prefrontal cortex (vmPFC), and the amygdala (Ruff & Fehr, 2014). Such experience-reward signals are important inputs to distinct neurons in OFC/vmPFC, amygdala, and striatum, which update and maintain the stimulus-value associations and expectations employed in judgment and decision-making (Balleine, Delgado, & Hikosaka, 2007; Berridge & Kringelbach, 2015; Wassum & Izquierdo, 2015). In contrast to rewards, aversive experiences (e.g. pain) are reliably associated with activity in aINS, dACC, and distinct neural populations in the amygdala (Pessiglione & Delgado, 2015). Importantly, aINS and dACC anchor the salience network which functions to increase attention to important changes in the environment and motivate avoidance of dangerous or noxious stimuli (Cunningham & Brosch, 2012; Shackman et al., 2011). Finally, the dorsolateral prefrontal cortex (dlPFC) supports goal maintenance and implementing specific goal-directed behaviors (Ruff & Fehr, 2014).
The brain areas described above are also implicated in social decision-making tasks (Ruff & Fehr, 2014). Though one study with neurological patients suggests that social and non-social value-based decision-making might be separable at the neural level (Besnard et al., 2016), the extant evidence is inconclusive. Since the most common techniques employed in human functional neuroimaging assess coordinated activity across large populations of neurons, it remains an open question whether social and non-social information is supported by the same neurons, or engage distinct parallel channels. For instance, vmPFC seems implicated in both social cognition and valuation, and this has sparked a debate about its primary functional role (Delgado et al., 2016). From a neuroeconomics perspective, the significance of vmPFC is in supporting the representation of the personal subjective value of stimuli with a single “common currency” (Ruff & Fehr, 2014). Others argue that the vmPFC instead primarily functions to support social cognition by maintaining representations of the self and close others (Delgado et al., 2016). An anatomically intact vmPFC is crucial for typical decision-making in tasks which require individuals to distinguish between different rewards and punishments with varying magnitudes and probabilities, such as the Iowa Gambling Task (Gläscher et al., 2012). Social contexts contain highly dynamic information and representations of other individuals. Thus, the vmPFC involvement in social cognition could arise in response to the processing demands required by such contexts. Moreover, mental state attribution, which is can be a necessary input to social decision-making, relies on a network of interconnected cortical regions including the posterior superior temporal sulcus (pSTS), temporoparietal junction (TPJ), medial prefrontal cortex (mPFC), posterior cingulate cortex (PCC), and precuneus (Decety & Lamm, 2007; Tremblay et al., 2017; Young, Cushman, Hauser, & Saxe, 2007). These regions supporting mental state attribution and dlPFC are coactivated specifically during social decision-making This functional coupling reflects the expected value of decisions and provides a crucial framework for social information such as mental state understanding to influence valuation processes (Decety & Yoder, 2017; Delgado et al., 2016).
Overall, though there are still open questions, cognitive neuroscience and neuroeconomics have provided a solid platform for investigating specific cases of social decision-making that are directly relevant to legal settings. First, research into moral cognition has identified the neural computations and networks important for assessing intentionality and harm, as well as how those systems interact to produce judgments of culpability and blameworthiness (Krueger & Hoffman, 2016). Second, the emerging field of neuroethics has begun to clarify the factors that contribute to antisocial behavior, including potential neuropsychological risk factors for future antisocial behavior (Gaudet, Kerkmans, Anderson, & Kiehl, 2016).
Moral values, empathy and justice motivation
People have moral values. They accept standards that allow their behavior to be judged as either right or wrong, good or evil, praiseworthy or blameworthy. Though particular norms by which moral actions are judged vary to some extent from individual to individual and across geographical locations (although some norms, such as not to kill, not to steal, and to honor one’s parents, are widespread and perhaps universal), all cultures demonstrate value judgments concerning human behavior. So important are social norms that in every culture studied to date people are willing to give up some of their own resources to punish individuals that violate social norms, even when they themselves are not directly affected (Henrich et al., 2006). Moreover, third parties evaluate moral wrongness and blameworthiness by taking into account both an action’s outcome and an agent’s intention (Buckholtz et al., 2008; Young, Scholz, & Saxe, 2011). In this way, individuals’ everyday moral judgments match the foundational legal concepts of mens rea and actus rea, where harmful intentions and harmful outcomes produce additive effects on condemnation. Thus, third-party judgments necessitate social decision-making capacities to integrate the value of particular outcomes and the understanding of the mental state of relevant agents, the latter relying on social cognitive abilities such as theory of mind.
Work across various academic disciplines has converged on the view that morality arises from the integration of both innate abilities which are shaped by natural selection and deliberative processes that interact with social environments and cultural exposure (Decety & Wheatley, 2015). Throughout most of human history individuals have lived in small groups which facilitated repeated interactions with other individuals (Baumard, André, & Sperber, 2013). Moreover, humans’ elaborated social cognitive abilities allow individuals to not only observe the behavior of others and predict future behavior, but also communicate that information to one another. Since individuals have some flexibility in selecting social partners, partner choice exerts strong pressure to at least appear as if one will likely cooperate and support reciprocal interaction (Baumard & Sheskin, 2015). This mutualistic perspective posits that the most efficient psychological mechanism to achieve a reputation as a cooperator is through a genuine moral concern which treats cooperation as intrinsically good (Baumard et al., 2013; Debove, Baumard, & André, 2017). In other words, it is evolutionarily beneficial for humans to genuinely prefer some kinds of moral outcomes because it helps to maintain cooperation.
Another theory of morality rests on the idea that humans are equipped with a moral disposition. This naturalistic view of a moral sense is originally grounded in the Humean idea that moral judgments arise from an immediate aversive reaction to perceived or imagined harm to victims (Hume, 1738). These actions are judged as immoral only after, and because of, the initial affective reaction. On that perspective, social emotions (e.g., empathy, guilt, shame) play a pivotal role in morality. Such social emotions contribute to fitness in facilitating caring for others and group cohesion.
Cognitive neuroscience of morality
Converging evidence from functional neuroimaging studies and neurological observations indicates that the same regions implicated in social decision-making play important specific roles in morality (Figure 1). Specifically, a set of interconnected regions encompassing the vmPFC, OFC, amygdala, TPJ, ACC, aINS, PCC, and dlPFC are reliably engaged across tasks which involve explicit or implicit evaluations of morally-laden stimuli, regardless of whether the outcome of an action affects the participants directly or another individual (Eres, Louis, & Molenberghs, 2017). Neuroscience investigations of third-party punishment judgments, such as those made by jurors (Buckholtz et al., 2008), suggest that intention understanding and harm perception rely on interconnected, but largely distinct neural systems (Krueger & Hoffman, 2016). As discussed above, representations of others’ mental states, beliefs, and intentions are supported by pSTS/TPJ, PCC/precuneus, and mPFC (Decety & Lamm, 2007; Young et al., 2007, 2011). These regions are foundational to incorporating intentionality into moral judgments. Conversely, perception of harm relies more on aINS, ACC, and the amygdala, core nodes of the salience network (Droutman, Bechara, & Read, 2015; Hesse et al., 2016), reflecting a neural mechanism for an actions’ outcome to influence decision-making. Finally, integrating harm and intent in order to determine an appropriate punishment relies on intact functioning of the central executive network, especially lateral parietal cortex and dlPFC (Buckholtz & Marois, 2012; Decety & Cowell, 2017; Krueger & Hoffman, 2016).
I. Development Divide
One of the most significant contributions of neuroscience to moral theory lies in understanding the developmental divide in moral behavior. Moral capacities are not evenly distributed across individuals or societies; they develop over time and are shaped by both biology and environment.
From a neuroscientific standpoint, moral development is closely tied to brain maturation. The prefrontal cortex, responsible for executive functions such as impulse control, planning, and ethical reasoning, continues developing into early adulthood. This explains why children and adolescents often display weaker moral restraint and greater susceptibility to peer pressure. Neuroscience thus supports long-standing psychological theories, such as those proposed by Jean Piaget and Lawrence Kohlberg, while grounding them in biological processes.
However, the development divide is not only age-related. Socioeconomic conditions, trauma, nutrition, and education significantly affect neural development. Children exposed to chronic stress or violence may experience alterations in brain regions involved in empathy and emotional regulation. This creates disparities in moral behavior not because of inherent moral inferiority, but due to unequal developmental conditions.
On a global scale, access to healthcare, education, and stable environments influences how moral norms are internalized. Neuroscience reveals that moral behavior cannot be separated from structural inequality. Thus, the “development divide” reflects both neurological maturation and broader social injustice, challenging simplistic explanations of morality as purely individual choice.
II. Opportunities and Challenges
Neuroscience offers unprecedented opportunities to deepen our understanding of moral behavior. One major opportunity lies in identifying the neural mechanisms of empathy and prosocial behavior. Studies show that mirror neuron systems and limbic structures enable individuals to feel others’ pain, providing a biological basis for compassion and altruism. Such findings can inform education, conflict resolution, and rehabilitation programs by emphasizing emotional engagement rather than punishment alone.
Another opportunity is in the field of moral pathology. Neuroscience helps explain why individuals with certain brain injuries or disorders—such as damage to the ventromedial prefrontal cortex—may exhibit antisocial or immoral behavior despite intact intelligence. This challenges traditional legal and moral assumptions about responsibility and intent.
Despite these advances, significant challenges remain. Neuroscience risks reductionism, the idea that morality can be fully explained by neural activity alone. Moral decisions are influenced by culture, language, religion, and social norms—factors that cannot be localized in a single brain region. Brain scans may show where activity occurs, but they cannot fully explain why certain moral values exist.
There is also the ethical challenge of interpretation. Neuroscientific data can be misused to label individuals or groups as morally deficient, reinforcing stigma or discrimination. Thus, while neuroscience opens new doors, it must be applied cautiously and ethically.
III. Strategies for Balanced Development
To responsibly use neuroscience in explaining moral behavior, balanced integration strategies are essential. Neuroscience should complement, not replace, philosophical ethics, psychology, and sociology.
One effective strategy is interdisciplinary research. Combining neuroscience with moral philosophy allows empirical findings to be interpreted within ethical frameworks such as virtue ethics, deontology, or consequentialism. For example, neuroscientific evidence showing the role of emotion in moral judgment challenges purely rationalist moral theories, but it does not eliminate the need for ethical reasoning.
Education is another critical strategy. Moral education informed by neuroscience can emphasize emotional literacy, empathy training, and self-regulation skills, especially during critical developmental periods. Programs that integrate mindfulness and social-emotional learning have shown positive effects on both brain development and moral conduct.
Finally, public communication must be carefully managed. Neuroscience should be presented as explaining tendencies and capacities, not determining destiny. Moral agency must remain a central concept, preserving the idea that individuals can reflect, learn, and change.
IV. Policy Frameworks and Historical Context
Historically, morality has been governed by religious codes, legal systems, and philosophical doctrines long before neuroscience existed. Ancient moral systems emphasized virtue, duty, or divine command, relying on social enforcement rather than biological understanding.
The emergence of neuroscience in the late 20th century introduced a new policy challenge: how to incorporate scientific insights into moral governance without undermining responsibility. Legal systems now grapple with neuroscientific evidence in courtrooms, particularly in cases involving criminal behavior, mental illness, or diminished capacity.
Policy frameworks must balance scientific insight with ethical safeguards. For example, while neuroscience can inform rehabilitation-focused justice systems, it should not justify invasive interventions such as compulsory brain modification. International human rights principles stress cognitive liberty and mental privacy, which must guide any policy application.
Historically, misapplications of biological explanations—such as eugenics—serve as cautionary tales. Neuroscience-informed policies must therefore be transparent, evidence-based, and grounded in respect for human dignity.
V. Case Studies in Integrated Development
Several real-world case studies demonstrate how neuroscience can be integrated into moral development initiatives. One example is rehabilitation programs for offenders that focus on impulse control and empathy training. Neuropsychological assessments help tailor interventions that reduce recidivism by addressing underlying cognitive deficits rather than relying solely on punishment.
Another case is early childhood intervention programs. Research shows that enriching environments during early brain development enhance emotional regulation and prosocial behavior. Programs that provide nutrition, parental support, and emotional learning reduce long-term antisocial outcomes, illustrating how neuroscience-informed policy can promote moral development.
In clinical settings, patients with brain injuries affecting moral judgment benefit from therapies that compensate for neurological limitations through structured decision-making support. These cases highlight that moral behavior emerges from an interaction between brain capacity and social scaffolding.
VI. Recommendations for Policy Prioritization
Based on neuroscientific insights, several policy recommendations emerge. First, governments should prioritize early childhood development, recognizing that moral capacities are shaped by neural growth during formative years. Investments in education, healthcare, and family support are effectively moral investments.
Second, criminal justice systems should integrate neuroscience-informed rehabilitation rather than focusing exclusively on retribution. Understanding the neural roots of impulsivity and aggression can lead to more humane and effective responses to crime.
Third, ethical oversight bodies should regulate the use of neuroscientific data to prevent misuse. Clear guidelines are needed to protect mental privacy and prevent discrimination based on neurological traits.
Finally, public education should promote neuroscience literacy to prevent deterministic interpretations of moral behavior. Citizens must understand that biology influences morality without eliminating choice or responsibility.
Conclusion
Neuroscience has profoundly reshaped our understanding of moral behavior by revealing the biological foundations of empathy, judgment, and self-control. It demonstrates that morality is not solely a product of abstract reasoning or cultural instruction but is deeply rooted in brain development and neural processes.
However, neuroscience cannot fully explain moral behavior in isolation. Moral values are shaped by history, culture, relationships, and conscious reflection. Reducing morality to neural activity risks oversimplification and ethical harm. Instead, neuroscience should be viewed as one vital component in a broader, interdisciplinary understanding of human morality.
When integrated responsibly, neuroscience offers powerful tools to reduce inequality, inform ethical policy, and promote moral development across societies. It explains how moral capacities arise, but the question of how we ought to live ultimately remains a shared human responsibility—guided by science, but grounded in ethical wisdom.
Over the past few decades, neuroscience has made great strides in understanding how the brain enables social behaviors, and how various disorders affecting the nervous system can inhibit an individual’s ability to engage in social interaction. While there is still a long way to go until the neuroscience of social behavior is fully understood, the insights already uncovered assist the development of medical treatments, and provide a platform for deeper investigation.